")
")
ロシュ分子生物学研究所のポスドク期間は2年と決められているので、筆者が着任して1年が過ぎると、前からいた3人のポスドク達は次々と別の研究施設へ移っていった。彼らの後半の1年間は、mRNAキャッピングという先端的研究に関わることができたこともあり、良い論文を連発していたので、次の職場へも喜んで迎えられていった。イスラエルから来ていたサラ(Sara Lavi)、オーストラリアからのジェリー(Gerry Both)はそれぞれ故国の国立研究所に就職し、インド人ポスドクのクリシュ(Muthukrishnan)はカンサス大学の助教ということで、それぞれ、良いポストを得て散って行った。そして、この3人の代りに入ってきたのは、のちに世界的に名を馳せることになるマリリン(Marylin Kozak:コザックルールの発見者)とナフム(Nahum Sonenberg:eIF4Eの発見者)の二人と、それから、少し遅れて、毛沢東が起こした文化大革命の混乱の中を、家族を中国に置いて、留学してきた中年の中国人ダショウ(Dashaw Wong)である。この3人はそれぞれの理由で印象深い。
この稿では、ナフムのキャップ結合因子eIF4Eの発見エピソードを中心に紹介し、あとの稿ではマリリンについて書き、ダショウについては、特に目覚ましい発見があったわけではないので紹介はしない。ただ、中国の文化大革命について一言いうと、それは1966年から10年間続き、その本質は、毛沢東が政治への復権を画して起こした権力闘争であり、民衆を暴動させて革新派を攻撃した。このため、中國の教育や科学技術は、その後20年以上も遅れることになったし、拷問や自殺で2000万人以上の命が失われたといわれる。民衆運動は反・高等教育へ向かい、科学教育を受けた人は迫害を受け、「学んだことを忘れることを脅迫された」とダショウはいう。そんな、境遇から、妻子を人質に取られて、留学してきたダショウであったから、アーロンから彼の教育を頼まれた私も大変だった。言葉の壁(英語がうまく通じない)があることは勿論、明治の頃のオジサンがタイムスリップして研究室に現れたような状況でもありーーそんな彼に生化学の基礎を教えることの難しさを想像してもらいたい。幸い、ダショウと私には共通してわかる「漢字」があり、とりわけ、私は中国の歴史が得意だったから、カタコト英語と筆談で緩やかではあるが、ダショウとの付き合いは順調に進んだ。
それにしても、何故、ダショウがーー世界最先端の研究を行っているロシュ分子生物学研究所へ来たのだろうか。私の推理だが、ユーデンフレンド所長は全米ジュウイッシュ協会の副会長だったから、Chinese Jewishの縁で、赤い中国から留学生を一人引き受けたのかもしれない。科学とか教育という言葉を発すると、殺されそうにもなる文化大革命の暗黒の時代に――中国の将来の科学をおもんばかって、研究者を留学させる政治指導者がいたことは、感心に値する。
キャップはどの様にしてmRNAの翻訳を促進するのかという質問
この質問については、この当時、m7Gと一価のプラスチャージが重要であること以外、全く不明であった。m7GpppN--の5′末端構造を持つmRNAは40Sのリボソームに優先的に結合し、タンパク合成の初期反応を速やかに進めることができる。それがどのようなメカニズムでそうなるのかは未知であった。この頃、アーロンも私も、セミナーに招ばれてmRNAキャッピングやタンパク合成の話をすることが多かったが、質問には、きまって“キャップはどの様にしてmRNAの翻訳を促進するのか”が出た。
答は、「mRNAは直線ではない。多分RNA自体が2次構造や3次構造を作って丸くなっているだろう。おまけに、タンパク合成の初期因子タンパクもそのmRNAに結合しているだろう。そのようなmRNA・タンパク複合体が、40Sリボソームに結合するためには、ーー多分、リボソームの穴にmRNAの糸を通すためには“糸の先に針のような構造”が必要でありーーそれがmRNA構造から突き出ているのではなかろうか、つまり、“mRNAの糸を通す:threading the string of RNA through the ribosomal hole by a stiff cap needle”でなかろうか」であった。そして、この答えは、この後に述べる、キャップ結合タンパクやタンパク合成の初期因子で裏付けされた実験データに矛盾してなく、ほぼ正解であろうと思われる。
これから10年ほど後には、キャップ結合タンパクには“mRNAの糸の2次構造をほぐす”活性が附随することが判り、「mRNAの5′UTRの構造変化によるタンパク合成の制御」とつながり、さらにその10年後には、「細胞の癌化のメカニズム解明」や「キャップ結合タンパクを標的とする癌創薬」まで、展開して行くのである。よく言われるーー基礎から応用ーーへの、基礎の第一歩、二歩がここにあった。
イスラエルからの帰還兵
1976年の秋だったろうか、若いイスラエル人ポスドクが着任してきた。ナフムという。彼は、第4次アラブ・イスラエル戦争の戦場から戻ってきた戦車兵士(タンクマン)であり、アーロンからは「教育して、実験ができるようにしてやってくれ」と頼まれていた。常時戦時体制下にあるイスラエルでは、兵役を終えた若い人は就学に関して海外留学などの優遇制度があるようである。私はナフムに付き添って1-2週間、日本の生化学で教わった教目を教え、特にキャップやRNAの化学については念入りに学習させた。ーーというのも、彼のテーマはキャップに結合するタンパクの解析であるからである。ナフムは、私と始めて出会った時、私が日本人であることに驚いたようであった。彼はダショウと違い、キャップに関する論文を読んでいたが、筆者Furuichiを、アルファベットのスペルから、日本人とは思わず、イタリア人かポーランド人と思っていたフシがある。実は、ナフムと筆者が出会った時を数年さかのぼる1972年に、イスラエルのテルアビブ空港で、3人の日本人テロリスト(日本赤軍:奥平・安田・岡本)により、機関銃の乱射や手榴弾破裂により、乗降客を含む26人が殺害され、70名を超える重軽傷者を出した事件があった。奥平・安田は自爆し、岡本は生き残り、現在もアラブの英雄として生きているが、この事件は、昨今、よく起こされる無差別自爆テロ事件のイスラム側のジハード(聖戦)のはしりとなった。このような事件が背景にあるので、アラブ・イスラエル戦線から戻ったナフムには日本人を恨む理由があった。「Do you know Kozo Okamoto?」が、彼が厳しい顔で、私に問うた最初の質問だった。言い訳になるが、正直、私は研究に没頭するあまり、日本赤軍や日本の若者が、何の関係もないアラブ・イスラエルの対立に首を突っ込んで、アラブ側に立ち、テロをやっていることをよく知らなかった。正直、申し訳なく思い、私は日本人として素直に謝った。一方、筆者は自分の興味から彼が戦車兵士としてどんな戦いをエジプト軍とやっていたのか尋ねた。「機関銃を打っていたのか、砲手だったのか、操縦士だったのか」などなど、である。ナフムは恥ずかしそうに答えたが、砂漠の中で、エジプトの戦車がどの方向に、どの距離に居るのか計算して砲手に伝える役だったそうである。
ヴィテックFillipowiczによるキャップ結合タンパクの発見
この当時、どのようなタンパク合成の初期因子がキャップを認識して結合するのかが、タンパク合成研究の喫緊の大きな問題だった。オチョアの研究室ではヴィテックが取り掛かった。彼はオチョア先生がこの問題に興味を持っていることをよくわかってなく、昼間はオチョア先生から言われたタンパク合成初期因子eIF2のリン酸化の研究を行い、夜にはビクビク・コッソリ、私とキャップ結合タンパクの研究をおこなった。彼はこっそりやっていたが、その心配は全く無かった。日本から、上代淑人先生が、NIHで開かれたタンパク合成のシンポジュームに来られ、オチョア先生はこの会の花形であった。私とマリリンKozakは、一緒にこのNIHの会に出席していたが、そこでの話題はキャップ結合タンパクであり、上代先生はオチョア先生に、キャップ結合タンパクの研究をやるように勧めていた。オチョア先生は私にウインクしながら「うちでもやっているよ」言っていられたが、それはヴィテックのことであり、お見通しだったのである。実際、ヴィテックを筆頭著者としたPNAS論文が、オチョア先生の主筆で完成した38。これは、RIMB研究所内の階を越え、部を越えて行われた共同研究成果であり、こんな具合に、RIMB研究所では、流行のキャップの研究を行う研究室が増えていた。この論文で嬉しかったのは、私は、恩師の上代先生のそのまた恩師のオチョア先生と共著で論文が発表できたことであり、「オチョア・スペインファミリー」の一員に加えてもらったことであった。論文では、ヴィテックは、私が作った放射性3Hや32Pでラベルしたキャップオリゴをアルテミア・エビ抽出液(brine shrimp extract)を細かく分画したタンパクと混合してニトロセルロース膜上に留まる複合体を作るかどうかを調べ、キャップに結合するタンパクがあることを突き止めていたが、それが何であるか突きとめることはできなかった。そんな中、彼もまた2年間のポスドク期間が来て、スイスのFMI研究所へ移っていった。オチョア先生の推薦と、最先端研究であるキャップ結合タンパクを報告するPNAS論文があれば、世界のどこの研究室へも就職できたと思われる。彼はその後、バーゼルに定住し、RNA研究でよい発見を遺すことになる。
Charlie Cantorのアドバイスが効いた、ナフムSonenbergによるキャップ結合タンパク(CBP)の発見
タンパク合成はその行程が、初期反応、ペプチド鎖の延長、終期反応の3ステップに分けられ、なかでも初期反応が重要であり、ここが注目の的であった。キャップはmRNAの先端にあり、しかも、それが無ければタンパク合成の初期反応がスタートしないことが判っていたから、キャップと結合するタンパク(cap binding protein: 以後CBPと略す)が、初期タンパク因子のなかにあるはずであり、それがヴィテックの先行する研究で確認できていた。そして、それがどんなタンパクであるのかを突き止めるのがナフムのミッションであり、ヴィテックが去ったあと、バトンはナフムに渡された。
ナフムと私は、ヴィテックの先行する研究によりーー存在することがわかったーー「幻のCBP」を、放射性キャップオリゴとのクロスリンクにより捕まえようとする実験にとりかかった。まず3H-MethylでラベルしたレオウイルスのmRNAを作り、先端にあるm7Gのリボースの2′-3′-cis-diol OHを過ヨード酸で酸化し、ジアルデヒドに変えるのである。つぎに、このmRNAを、小麦胚芽抽出液(wheat germ extract)から抽出したタンパク合成の初期因子や、リボソームからはがしたタンパク質と混和して、短時間インキュベートする。すると、キャップにゆるく結合したCBPの中に、m7Gのアルデヒドと結合できるリジンなどのアミノ基があれば、反応して -CH=N- なるシッフ塩基(Schiff base)を形成してキャップとCBPタンパク質を安定した共有結合でつなぐことはずである。幸いなことに、mRNAはキャップを酸化しても40Sリボソームと結合するので、タンパク合成の初期行程はクリアしていることがわかった。
ただ、ここでできるシッフ塩基結合C=Nは不安定なので、還元してC-Nにしないと加水分解して元に戻ってしまう。そこで、C=Nを安定化する還元剤が必要であり、筆者はNaBH4(ナトリューム・ボロン4水素)という化合物を使うことをナフムに勧めた。そして、その一回目の実験の反応物をポリアクリルアミドゲル・電気泳動(PAGE)で分析してみたところ、残念ながら、3Hでラベルされるタンパク質は検出されなかった。ーーということで、二人はガッカリした。筆者はあきらめが早い(特に、この場合は、「早すぎた」と反省している)。「この実験ではだめだ、何か別のことを考えよう」とナフムを残して、議論の場から去ってしまった。ところが、ナフムは中々あきらめない、そしてそれがよかった、ーーその粘りが、彼の運命を変えたともいえる。彼は、アーロンに頼んでシッフ塩基の安定化反応について知る化学者を紹介してくれるよう頼んだ。名前が挙がったのは、コロンビア大学教授のチャーリーCantor(Charles Cantor)である。チャーリーからは、即座に、我々が最初に使ったNaBH4よりは還元力の強いNaBH3CNを使えというアドバイスが帰ってきた。これは最高のアドバイスだった。二回目の実験では、いくつかのタンパク質がクロスリンクして見えるようになった。この時以来、このようなクロスリンクの実験ではNaBH3CNが使われるが、その成功例を早い段階で示したナフムの粘り強い挑戦と、チャーリーの貴重なアドバイスの意義は大きい。とても異例なことであるが、この実験結果を報じたPNAS論文39のDiscussionの文中に「C.Cantorのアドバイスに従ってやった」と書かれている。実際、チャーリーのアドバイスが無かったらCBPの発見は1~2年遅れたであろう。
RNA研究を進展させたCBP(eIF4E)の同定
このmRNAのクロスリンク実験により、はじめのうちは数本のタンパクが「幻のCBP」の候補として浮かび上がっていたが、第5話で述べたm7GDPを阻害剤として使うBaglioniの方法を組み合わせることにより、2万4千ダルトンの分子量を持つ1個のタンパク質がタンパク合成に必要なキャップ結合タンパク(CBP)であることが判った39,40。写真はこの頃、ナフムとアーロンが実験室で、CBP発見の論文をPNAS誌へ送るための校正をやっている様子を撮ったものであり、撮影者は筆者である。私はこの論文の共著者になることを固辞した。ナフムの輝かしい門出の論文であり、私の寄与は小さいと思ったからである。ただ、この写真に写っている二人が、10年後、20年後にMCB誌の初代と2代目のEditor-in-Chiefになろうとは思いもよらないことであった(図1)。

図1: 実験室で24K CBP発見のPNAS論文を校正中のNahum Sonenberg(左)とAaron Shatkin(右)
この24K CBPは、その後ナフムによって、m7Gppアフィニティーカラムクロマトグラフィーで精製され、eIF4Eと名付けられた。eIF4E はタンパク合成実験系に加えると、細胞のmRNAなどキャップをもつmRNAのタンパク合成を促進するが、ポリオウイルスやC型肝炎ウイルスのmRNAなどキャップを持たないウイルスmRNAのタンパク合成には促進効果を示さない。これがどのように働くかについて、ナフムが紹介している24K CBPのタンパク合成初期における生物学的役割を図2で示す41。端的に言えば、eIF4Eは初期因子複合体の中にあって、他の種々の機能を持つ因子を引き連れて正確にmRNAの5′末端へ結合し、図2に示すように2次構造を解きほぐしたりして、40SリボソームがmRNA上を動き易くするのである。このため、小さなeIF4E(24K)はRNAヘリカーゼ活性を持つeIF4A(47K)と、この二つのタンパクを載せるeIF4G(220K)の3つからなる複合体の中に収まっていることが判った。
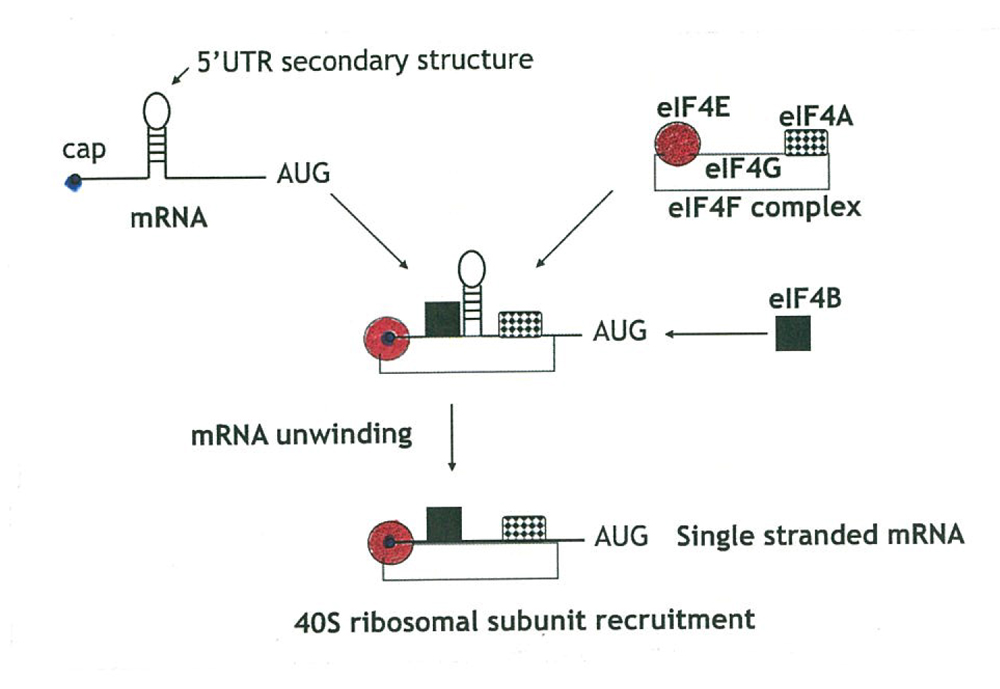
図2: タンパク合成におけるキャップ結合タンパクeIF4Eの役割(引用文献41)から改変: eIF4E の結合に伴いmRNAの2次構造がほどけることに注目
キャップ結合タンパクeIF4Eは、mRNAの翻訳を介してがんの発生につながることが判り、そのシグナリング4E-BPと共に、がん創薬の標的になっている
1990年のある日、日本に帰っていた私のところへ、カナダMcGill大の教授として独立した研究室を持ち、CBPの研究を続けていたナフムから驚きのメールが届いた。CBPがプロトオンコタンパク(がん促進タンパク)であることを示すデータが出たというメールであった。癌はDNA変異による病気といわれて久しいが、ナフムのデータは、正常なCBPタンパクを正常な細胞中で過剰発現してやると、細胞は癌化するという驚きの大発見である。この時、アーロン、ナフム、それに筆者の3人は、それぞれ別れて、異なる研究へ挑戦していた。ちなみにアーロンはCABMという大きな研究所を、ラトガース大構内のワックスマン研究所に隣接して創り、日々、研究所の経営に追われて忙しく、キャップ研究は続けてはいるものの、メジャーな仕事ではなくなっていた。一方、筆者といえば、帰国して以来、鎌倉にある日本ロシュ研究所で、内皮細胞が分泌するエンドセリンという血圧調整ペプチドに対する創薬研究や、ビタミンCをバイオ技術で作る研究や、ロシュ主催の国際創薬シンポジュームを開催する仕事に追われーーすでにキャップ研究からは卒業していたのであったが、このニュースには驚いた。そして、この発見を記述した論文はすでにNature誌に採択された42ということを聞き、この発見をアーロンと一緒に大いに喜んだのである。ナフムの発見と言うか、Sonenberg研のポスドクLazaris-Karatzas博士の発見は、CBPの最重要成分であるeIF4Eタンパクを多く発現すると、普段は強固な2次構造の5′UTR(mRNAの5′領域にあってタンパク質に翻訳されない部分)のせいで、翻訳されなかった発がんタンパクのmRNAが、(CBP中のRNAヘリカーゼeIF4Aによって、図2のように、2次構造が解かれて)翻訳されるため、発がんタンパク(例えばmyc)が多く作られて、癌化するというのである42,43。これはどこから見ても正解である。
ただ、「キャップはタンパク合成を促進する」と簡単に言い切れない問題が、ここにはある。mRNAの2次構造には色々あり、容易に翻訳されるmRNAと「そうでないmRNA」があり、細胞の分化や増殖のための重要なシグナルとなるタンパクの場合は、ことさら大事で、mRNAは細胞中に常在していても、フリーのeIF4Eの量が少なければ、タンパクは合成されない。だから、「mRNAがたくさんあれば、その方向に細胞は動くわけではなく、遺伝子発現は翻訳レベルでも調節されている」のである。翻訳レベルの調節ということは、かねてより、討論されてきたが、スッキリした答えは得られていなかった。しかし、ここで確度の高い答えが得られた。
細胞内にフリーでいるeIF4Eの量は、また、その後、ナフムの研究室で見つけられた4E-BP(4E-Binding Proteinから名前が取られている)というタンパクにより調節されていることが判った。4E-BPがeIF4Eに結合すると、eIF4Eは(eIF-4Gと結合できなく)mRNA上のキャップと結合できなくなり、働きは抑えられる。このことから、4E-BPは「がん抑制タンパク(サプレッサー)」ともいわれている。
しかしながら、4E-BPもまた、はリン酸化され、eIF4Eに結合できなくなるという具合になっていて、幾重にも、巧妙にシグナルの調節がかけられているから複雑である。ただ、この原理に基づき、現在、がん創薬へ向けて臨床試験が行われている。たとえば、イーライリリー社は、アンチセンスDNAを用いて、eIF4EのmRNAを壊す抗がん剤DNAを開発中であり(2008年)、また4E-BPのリン酸化を抑えるタイプの抗がん剤候補(たとえばラパマイシン)も検討中である。
キャップ結合タンパクの発見を皮切りに、ナフムとMcGill大のポスドク陣(日本からの留学生を多く含む)の活躍は素晴らしく、次々と新しい発見を重ねていった。この稿では、「mRNAと発がんメカニズム」に焦点を当てて、彼の発見のいくつかを紹介したが、いささか難解だったかもしれない。実は、ウイルスの感染戦略に関しても、キャップ結合タンパクに絡む、ストレートに面白い発見もあるので、次稿で紹介したいと思っている。
四方を見渡して、砂山以外は何も見えない。そんな光景の中に、第4次アラブ・イスラエル戦争で、タンクマンとして、標的を捜して戦っているナフムが見える。また、それがあるから、その後の研究現場でも、先の見えないRNA研究の中で闘ってきた彼の40年の活躍が想像できる。図1の、若い折の写真に合わせて、比較的最近、鎌倉へ筆者を訪ねて来てくれた折の、彼の写真を載せることにする(図3)。

図3: 鎌倉長谷寺にて、Nahum Sonenberg 博士と筆者
2度脱帽した、天才科学者チャーリー・カンター博士
チャーリーがナフムに天啓ともいうべき良いアドバイスを与えたことを先に書いた。チャーリーがアーロンの友人であることは知っていたが、1976年当時、リボソームRNAの研究をやっているという噂であったが、どのような発見をしたかについては全く知らなかった。ある夕べ、誘われて、アーロンとFort Leeにある日本食レストランへ出かけた。そこで、カウンターで寿司をつまんで飲んでいる小柄な男と出会い、紹介してもらった。それがチャーリーCantorだった。彼は日本食が大好きで、「よく一人で食べにくる」のだと言っていた。確かに、コロンビア大からはハドソン川をはさんで向かい側になり、遠くはない。この夕べは、アーロンがチャーリーへのお礼のために設けた一席だったのだろうーー、ただし、店は、チャーリーが指定したということであったろうかーー。日本食の人気が高まった現在はともかく、ニューヨーカーといえども、70年代に寿司が好きなヒトは稀であり、チャーリーは時代を先駆けるひとだ。筆者はナフムへのNaBH3CNのアドバイスに感心したことを述べたが、「なんてことはないよ」ということで、軽く一蹴されてしまった。また別の機会に、彼が「質量分析技術でRNAやDNAの塩基配列が効率よく決定できる」という内容の講演をしているのを聞いたことがあるが、私には全く理解できなかった。チャーリーは、奇才の人であり「10年も20年も、先が見える科学者ではなかろうか」という気がしていた。そうだ、のちに作られた映画『バック・トゥ・ザ・フューチャー』に登場するエメット・ブラウン博士のような印象を彼に持ったのである。
それから35年後、「ああっ、またやられた」とチャーリーに脱帽する大きな発見に出会った――このエッセイは、RNAに関する発見を中心に書いているが、ここではちょっと脱線して、天才チャーリーの思考と発見をフォローしたい。ことは、「胎児のダウン症の診断」に関することである。ダウン症は、21番染色体を通常の2本ではなく3本持つ胎児に起こる病気であり、高年齢女性の妊娠の際に頻度が高まる、「トリソミー細胞」が原因である。トリソミーは、21番染色体だけでなく、13番や18番染色体にも起るが、21番トリソミーが最も多い。このための出生前診断としては、従来、妊娠数か月後に、子宮に長い針を刺して羊水中に浮遊している胎児細胞を取りだし、細胞を培養後、細胞中の21番染色体の数(2本か3本か)をカウントする方法が行われてきた。この方法は、しかしながら、胎児への障害リスクがあるため、妊娠1~2か月の初期に妊婦の血液検査により診断ができないものかと思案がなされていた。金沢医科大学の研究チームは、胎児由来の有核赤血球が母親の血液1 cc中に1~2個現れることを発見し、この現象をダウン症の出生前診断に使うことを考えた。これは素晴らしい発見であるが応用が難しい。血中には、母親由来の無数の赤血球やその他の細胞が存在するが、そこから胎児の有核赤血球細胞を選別して拾い出すのである。ロボットと顕微鏡と、電動ピペットや細胞染色を駆使した出生前診断技術の開発研究が行われ、筆者はそのアドバイザーだった。一方、チャーリーは、母親の血液中に現れる21番染色体由来のDNA断片を、母親由来のDNAごと、次世代DNAシークエンサーで解析する方法を採用した。このために、彼は資金を集めてSequenom社というベンチャー企業を立ち上げている。
筆者は、DNAシークエンサーによる解析でMutationを捜すのはともかく、胎児の21番由来のDNAがほんの少量(1.5倍)多いーーそのわずかな差をどうして測定しーー、それによって重大な診断が下せるのか、全く理解できなかった。北陸の方法は手間がかかるが、結果が目に見えるのであり、チャーリーの方法では数字が出るだけであるーー。さて、長い話を短くして、結論を言うと、完全に彼の、Sequenom社の、勝ちだった。量的に僅かの差であっても、検体当たり数百回~数千回読めば差は百倍から数千倍に増幅されるのである。これには、脱帽であった。
筆者は、完敗を認め(ーーもっとも戦っていたわけでもないがーー)チャーリーに頼んで、金沢へ来てもらった。そしてSequenom社の無侵襲的出生前遺伝学的検査について講演を、お願いし、北陸のグループと共同で何かできないか考えてもらった。その時の写真を図4に示す。筆者は、槍・刀を巧みに駆使して創り上げた北陸の診断法が、チャーリーの機関銃を使った戦術に負けた気がして、アドバイザーとしてしばらく眠れない日が続いた。いまや、Sequenom社は世界のトリソミー症の診断にかけてはWorld Leader である、そして、チャーリーは大金持ちであろう。加えて、毎年、数万人以上の妊婦がダウン症診断の恩恵をうけているはずであり、素晴らしい。
(了)

図4: 金沢訪問のSequenon 社最高顧問Charles Cantor博士(右)と筆者(左)
< 第一回 | 第二回 | 第三回 | 第四回 | 第五回 | 第六回 | 第七回 | 第八回 | 第九回 | 第十回 | 第十一回 | 第十二回 | >
References
1. Miura K, Watanabe K, Sugiura M, Shatkin AJ.
The 5′-terminal nucleotide sequences of the double-stranded RNA of human reovirus.
Proc Natl Acad Sci U S A. 1974 Oct;71(10):3979-3983.
PMID: 4530278
2. Furuichi Y.
"Methylation-coupled" transcription by virus-associated transcriptase of cytoplasmic polyhedrosis virus containing double-stranded RNA.
Nucleic Acids Res. 1974 Jun;1(6):809-822.
PMID: 10793759
3. Smith AE.
Modified nucleotides in messenger RNA?
Nature 1974 Aug;9 250(5466) 461.
PDF (237K)
4. Furuichi Y, Miura KI.
Identity of the 3′-terminal sequences in ten genome segments of silkworm cytoplasmic polyhedrosis virus.
Virology. 1973 Oct;55(2):418-425.
PMID: 4742779
5. Furuichi Y.
Discovery of m(7)G-cap in eukaryotic mRNAs.
Proc Jpn Acad Ser B Phys Biol Sci. 2015;91(8):394-409.
PMID: 26460318
6. Furuichi Y, Miura K.
A blocked structure at the 5′ terminus of mRNA from cytoplasmic polyhedrosis virus.
Nature. 1975 Jan 31;253(5490):374-375.
PMID: 163011
PDF (414K)
7. Furuichi Y, Morgan M, Muthukrishnan S, Shatkin AJ.
Reovirus messenger RNA contains a methylated, blocked 5′-terminal structure: m-7G(5′)ppp(5′)G-MpCp-.
Proc Natl Acad Sci U S A. 1975 Jan;72(1):362-366.
PMID: 1054511
8. Wei CM, Moss B.
Methylated nucleotides block 5'-terminus of vaccinia virus messenger RNA.
Proc Natl Acad Sci U S A. 1975 Jan;72(1):318-322.
PMID: 164018
9. Adams JM, Cory S.
Modified nucleosides and bizarre 5'-termini in mouse myeloma mRNA.
Nature. 1975 May 1;255(5503):28-33.
PMID: 1128665
PDF (1.6M)
10. J. Darnell H. Lodish D. Baltimore
Molecular Cell Biology
Scientific American Books, W. H. Freeman and Company. (1986)
11. James E. Darnell.
RNA: Life's Indispensable Molecule
Cold Spring Harbor Laboratory Press. ISBN: 978-1-936113-19-4.
CSH Press
Amazon.co.jp
12. Furuichi Y, Morgan M, Shatkin AJ, Jelinek W, Salditt-Georgieff M, Darnell JE.
Proc Natl Acad Sci U S A. (1975) 72(5):1904-1908.
PMID: 1057180
13. Furuichi Y, Muthukrishnan S, Tomasz J, Shatkin AJ.
J Biol Chem. (1976) 251(16):5043-5053.
PMID: 821947
14. Abraham G, Rhodes DP, Banerjee AK.
Cell. (1975) 5(1):51-58.
PMID: 165900
15. 古市 泰宏
蛋白質・核酸・酵素 (1977) 22(10), 1201-1204.
16. Sommer S, Salditt-Georgieff M, Bachenheimer S, Darnell JE, Furuichi Y, Morgan M, Shatkin AJ.
Nucleic Acids Res. 1976 Mar;3(3):749-765.
PMID: 1272797
17. Amiya K. Banerjee
Proc Natl Acad Sci U S A. (2012) 109(46):18629-18630.
18. Furuichi Y, Shatkin AJ, Stavnezer E, Bishop JM.
Nature. 1975 Oct 16;257(5527):618-620.
PMID: 170541 PDF
19. Gelinas RE, Roberts RJ.
Cell. 1977 Jul;11(3):533-544.
PMID: 884734
20. Thomas M, White RL, Davis RW.
Proc Natl Acad Sci U S A. 1976 Jul;73(7):2294-2298.
PMID: 781674
21. Berget SM, Moore C, Sharp PA.
Proc Natl Acad Sci U S A. 1977 Aug;74(8):3171-3175.
PMID: 269380
22. 222.
Proc Natl Acad Sci U S A. 1977 Aug;74(8):3171-3175.
PMID: 269380
22. 222.
Proc Natl Acad Sci U S A. 1977 Aug;74(8):3171-3175.
PMID: 269380
23. Plotch SJ, Krug RM.
J Virol. 1977 Jan;21(1):24-34.
PMID: 833924
24. Bouloy M, Plotch SJ, Krug RM.
Proc Natl Acad Sci U S A. 1978 Oct;75(10):4886-90.
PMID: 283399
25. Furuichi Y, Shatkin AJ.
Proc Natl Acad Sci U S A. 1976 Oct;73(10):3448-52.
PMID: 185613
26. Bouloy M, Morgan MA, Shatkin AJ, Krug RM.
J Virol. 1979 Dec;32(3):895-904.
PMID: 513205
27. Furuichi Y.
Review:Caps on Eukaryotic mRNAs. In: eLS. John Wiley & Sons, Ltd: Chichester.
DOI: 10.1002/9780470015902.a0000891.pub3
28. Krug R.M. and Fodor E.
Webster-Textbook of Influenza
Wiley
Amazon
29. Urushibara T, Furuichi Y, Nishimura C, Miura K.
FEBS Lett. 1975 Jan 1;49(3):385-9.
PMID: 162886
30. Shatkin AJ.
Proc Natl Acad Sci U S A. 1974 Aug;71(8):3204-7.
PMID: 4528644
31. Hickey ED, Weber LA, Baglioni C.
Nature. 1976 May 6;261(5555):71-2.
PMID: 944859
32. Enigma Variations’ of mammalian messenger RNA. (News & Views)
Nature. 1975 May 1;255(5503):9
PDF
33. Muthukrishnan S, Both GW, Furuichi Y, Shatkin AJ.
Nature. 1975 May 1;255(5503):33-7.
PMID: 165427
34. Both GW, Banerjee AK, Shatkin AJ.
Proc Natl Acad Sci U S A. 1975 Mar;72(3):1189-93.
PMID: 165487
35. Both GW, Furuichi Y, Muthukrishnan S, Shatkin AJ.
Cell. 1975 Oct;6(2):185-95.
PMID: 1182800
36. Furuichi Y.
Proc Natl Acad Sci U S A. 1978 Mar;75(3):1086-90.
PMID: 349555
37. Furuichi Y, LaFiandra A, Shatkin AJ.
Nature. 1977 Mar 17;266(5599):235-9.
PMID: 557727
38. Filipowicz W, Furuichi Y, Sierra JM, Muthukrishnan S, Shatkin AJ, Ochoa S.
Proc Natl Acad Sci U S A. 1976 May;73(5):1559-63.
PMID: 1064023
39. Sonenberg N, Shatkin AJ.
Proc Natl Acad Sci U S A. 1977 Oct;74(10):4288-92.
PMID: 270673
40. Sonenberg N, Morgan MA, Merrick WC, Shatkin AJ.
Proc Natl Acad Sci U S A. 1978 Oct;75(10):4843-7.
PMID: 217002
41. Nahum Sonenberg
Biochem Cell Biol. 2008 Apr;86(2):178-83. doi: 10.1139/O08-034.
PMID: 18443631
42. Lazaris-Karatzas A, Montine K.S. Sonenberg N.
Nature. 1990 Jun 7;345(6275):544-7.
PMID: 2348862
43. Lazaris-Karatzas A. Sonenberg N.
Mol Cell Biol. 1992 Mar;12(3):1234-8.
PMID: 1532049